خلاصه ی محتوای علمی چهارمین نشست کارگروه نوروفیزیولوژی شاخه دانشجویی نقشه برداری مغز ایران
یکی از پیچیده ترین سیستم های عملکردی بدن، دستگاه عصبی می باشد. بنابراین برای درک اعمال اساسی این سیستم ابتدا باید کلیات آن را در اینجا مورد بحث قرار دهیم.
سازماندهی پایه ای دستگاه عصبی
یکی از پیچیده ترین سیستم های عملکردی بدن، دستگاه عصبی می باشد. بنابراین برای درک اعمال اساسی این سیستم ابتدا باید کلیات آن را در اینجا مورد بحث قرار دهیم. واحد عملکردی سیستم عصبی یا به عبارتی سلولهای موجود در دستگاه اعصاب، نورون (Neuron) نامیده می شود. هر نورون از یک بخش گیرنده اطلاعات (دندریت)، یک جسم سلولی (Soma) و یک بخش خروجی (آكسون) تشکیل شده است.
دستگاه عصبی از طریق گیرنده های حسی، اطلاعات را از محیط دریافت می کند (به عنوان مثال، گیرنده های بینایی در چشم ها، گیرنده های شنوایی در گوش ها، گیرنده های لمسی روی سطح بدن با سایر گیرنده های بدن)، سپس آنها را برای پردازش به مناطق جمع بندی کننده ی مغز می فرستد و سرانجام اطلاعات حاصله برای تنظیم فعالیت های مختلف بدن از طریق اعصاب حرکتی به اندام های محیطی فرستاده می شوند. اعمال حرکتی اعصاب به سه طریق می باشد و انقباض عضلات اسکلتی، ۲) انقباض عضلات صاف الحشاء داخلی و ۳) ترشح مواد شیمیایی فعال از غدد درون ریز و برون ریز در بسیاری از قسمت های بدن.
حافظه
حافله به روند ذخیره اطلاعات در سیستم عصبی، حافظه (۱۳۹۱) گفته می شود. ذخیره ی اطلاعات عمدتا در قشر مخ صورت می گیرد ولی مناطق قاعده ای مغز و نخاع نیز می توانند مقدار اندکی از اطلاعات را ذخیره کنند. حافظه از اعمال سیناپس ها می باشد. هر بار که نوع خاصی از سیگنال های عصبی از یک توالی سیناپسی عبور می کند، بعدا این سيناپس ها برای انتقال همان نوع سیگنال توانمندتر می شوند. این روند، تسهیل نام دارد. پس از آنکه سیگنال های حسی چندین بار از سیناپس ها عبور کردند، سیناپس ها به حدی تسهیل می شوند که حتی سیگنال هایی که بدون وجود محرک حسی ورودی، در خود مغز تولید شده اند نیز می توانند از طریق همان توالی مشابه سیناپسی منتقل شوند.
سطوح عملکردی دستگاه عصبی
دستگاه عصبی را از نظر عملکردی به سه بخش تقسیم می کنند: ۱) سطح نخاعی، ۲) سطح مغزی تحتانی یا سطح زیرقشری و ۳ سطح مغزی فوقانی با سطح قشری. نخاع قادر است بسیاری از حرکات رفلکسی مانند حرکات راه رفتن و پس کشیدن اندام ها از محرک های دردزا را سازماندهی کند. البته مراکز مغزی می توانند با ارسال پیام هایی به مراکز نخاعی، عملکردهای آنها را تعدیل نمایند. سطح مغزی تحتانی شامل بصل النخاع، پل مغزی، مزانسفال، هیپوتالاموس، تالاموس، مخچه و عقده های قاعده ای می شود. این مراکز بسیاری از فعالیت های ناخودآگاه بدن را کنترل می کنند که در ارتباط با آنها در بخش های بعدی صحبت خواهد شد. در سطح قشری، پردازش افکار و ذخیره اطلاعات و همچنین مقایسه اطلاعات ذخیره شده با اطلاعات جدید رسیده صورت می گیرد. البته قشر مغز برای عملکرد خود به سطوح زیر قشری نیاز دارد و به تنهایی قادر به فعالیت نیست.
سناپیس های دستگاه عصبی مرکزی
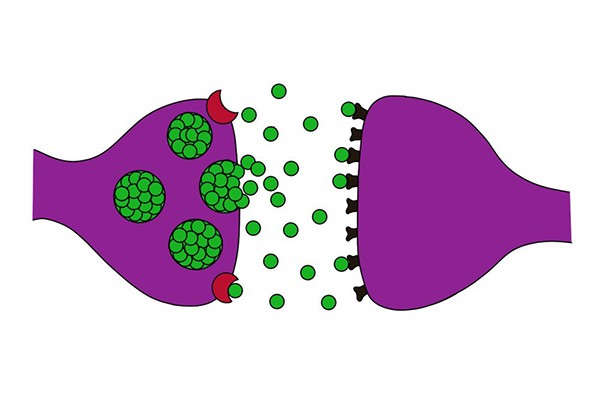
به طور کلی به محل اتصال یک نورون با نورون دیگر، سیناپس اطلاق می گردد. سیناپس ها به دو نوع اشیمیایی و لکتریکی تقسیم می شوند. تقریبا تمام سیناپس های دستگاه عصبی مرکزی از نوع سیناپس های شیمیایی هستند. در این سیناپس ها، اولین نورون یک ماده شیمیایی در سیناپس انتهای عصبی (پایانه پیش سیناپسی) ترشح می کند که ناقل عصبی (نوروترانسمیتر) نام دارد. این ناقل به نوبه ی خود بد پروتئین های گیرنده ی موجود در غشا نورون بعدی (پایانه پس سیناپسی) اثر می کند و سبب تحریک یا مهار نورون یا تغییر حساسیت آن می شود. سیناپس های الکتریکی که بیشتر در سلول های عضله ی صاف و قلبی وجود دارند، امکان انتشار جریان های الکتریکی از یک سلول به سلول دیگر را از طریق اتصالات شکافی فراهم می آورند. یک تفاوت عمده سیناپس الکتریکی با شیمیایی این است که سیناپس الکتریکی می تواند سیگنال-ها را در دو جهت هدایت کند اما سیناپس شیمیایی، سیگنال ها را فقط در یک جهت از نورون پیش سیناپسی به نورون پس سیناپسی ارسال می کند.
به پایانه پیش سیناپسی که معمولا روی دندریت و نا حدی روی جسم سلولی نورون پس سیناپسی، سیناپس ایجاد می کند، گره پایانه ای، دکمه یا پایک انتهایی با گره سیناپسی اطلاق می شود. بین این پایانه و پایانه پس سیناپسی یک شکاف سیناپسی قرار دارد که ماده ناقل عصبی به داخل این فضا آزاد می شود. در داخل پایانه پیش سیناپسی، وزیکول های ماده ناقل و میتوکندری وجود دارد وزیکول ها حاوی ماده ناقل عصبی بوده و میتوکندری ها آدنوزین تری فسفات (ATP) را می-سازند که انرژی لازم برای سنتز مواد ناقل جدید را فراهم می کند.
حال ببینیم که چه عاملی سبب می شود تا وزیکول های حاوی ماده میانجی، محتویات خود را به داخل شکاف سیناپسی رها کنند. زمانی که پتانسیل عمل ایجاد شده در یک نورون به انتهای آکسون آن نورون (پایانه پیش سیناپسی) می رسد، موجب می شود تا کانال های کلسیمی وابسته به ولتاژ در آن قسمت فعال شوند. در این حالت مقادیر زیادی کلسیم وارد پایانه پیش سیناپسی می گردد. این یون های کلسیم به مولکول های پروتئینی خاصی در سطح داخلی غشای پیش سیناپسی متصل می شوند که محل های آزادسازی نام دارند اتصال کلسیم به این محل ها لازمه آزاد شدن وزیکول های حاوی ماده میانجی به داخل شکاف سیناپسی می باشد به طوری که مقدار ماده ناقل رها شده به طور مستقیم با تعداد یون های کلسیمی که وارد پایانه پیش سیناپسی می شوند، ارتباط دارد.
ماده ناقل پس از آزاد شدن از پایانه پیش سینایسی به گیرنده های پروتئینی اختصاصی خود در غشا پایانه پس سیناپسی متصل می شود. این گیرنده ها یک بخش متصل شونده به ناقل عصبی دارند که در سطح بیرونی غشاء نورون پس سیناپسی قرار گرفته و یک بخش میان غشایی دارند که كل عرض غشاء را طی می کند. این می تواند یک کانال یونی باشد با یک فعال کننده ی پیام رسان ثانویه داخل سلولی باشد. برخی از موارد گیرنده خاص خود به طور مستقیم سبب باز شدن یک کانال یونی می گردند. اگر کانالی که باز شده است. کانال سدیمی باشد، مقادیر زیادی یون سدیم وارد سلول شده و سبب تحریک سلول یا دپلاریزاسیون می شوند. اگر کانال های پتاسیم و یا کانال های آنیونی باز شوند، خروج یون پتاسیم از سلول و ورود آنیون ها (مانند کلر) به داخل سلول سبب هیپرپلاریزاسیون (افزایش بار منفی) سلول و مهار نورون پس سیناپسی خواهند شد. مواد ناقلی که با اتصال به گیرنده های خود بر غشا نورون پس سیناپسی، سیستم پیام رسان ثانويه داخل سلولی را فعال می کنند، برخلاف موادی که بر کانال های یونی اثر دارند، سبب تغییرات طولانی مدت از چند ثانیه تا چند ماه پس از حذف ماده ناقل اولیه در نورون می کردند. یکی از معمول ترین انواع پیام رسان ثانویه، پروتئین های G هستند پروتئین های G معمولا در سطح داخلی غشاء به گیرنده پروتئینی ماده ناقل متصل می شوند. این پروتئین ها از سه جزء تشکیل شده اند: یک جز الفا که قسمت فعال کننده پروتئین می باشد و اجزای بتا و گاما که به جزء آلفا و به درون غشای سلولی مجاور گیرنده پروتئینی متصل هستند. اتصال ماده ناقل عصبی به گیرنده پروتئینی سبب جداشدن زیر واحدآلفا از زیر واحدهای بتا و گاما می گردد. جزء القای فعال شده می تواند موجب باز شدن کانال های یونی، فعال کردن آنزیم ادنیلیل سیکلاز و گوانیلیل سیکلاز و تولید cAMP (آدنوزین مونوفسفات حلقوی) و cGMP (گوانوزین مونوفسفات حلقوی)، فعال کردن یک یا چند آنزیم درون سلولی و فعال کردن رونویسی از برخی ژن ها گردد.
گیرنده های تحریکی و مهاری غشاء پس سیناپسی اگر اتصال یک ماده ناقل عصبی به گیرنده اختصاصی خود سبب شود تا کانال های سدیمی باز شوند، هدایت از کانال های پتاسیمی یا کلری متوقف شود که این امر سبب دپلاریزاسیون نورون می گردد و با تغییراتی را در متابولیسم داخل نورون پس سیناپسی ایجاد کند که سبب تحریک فعالیت سلول گشته، تعداد گیرنده های غشایی تحریکی را افزایش داده یا تعداد گیرنده های مهاری را در غشاء کاهش دهد. این ماده یک ماده تحریکی می باشد. برعکس، یک ماده مهاری با اتصال به گیرنده خود می تواند سبب باز کردن کانال های کلری و ورود یون کلر به داخل سلول (هیپرپلاریزاسیون) و یا افزایش هدایت یون های پتاسیم به خارج از نورون گردد و همچنین می تواند سبب فعال شدن گیرنده های آنزیمی ای گردد که فعالیت سلول را مهار می کنند به طور کلی هر عاملی که سبب شود پتانسیل استراحت غشاء به صفر نزدیک تر شود، یک عامل تحریکی بوده و اگر پتانسیل استراحت غشاء را به سمت منفی سوق دهد، یک عامل مهاری می باشد.
اگر گیرنده )رسپتور) نوروترانسمیتر، خود یک کانال یونی باشد، به آن رسپتور آینوتروپیک گفته می شود، در صورتی که گیرنده هایی که از طریق سیستم های پیام رسان ثانویه عمل می کنند، رسپتورهای متابوتروپیک نامیده می شوند.
نوروترانسمیترها با میانجی های عصبی
به طور کلی میانجی های عصبی آزاد شده از نورون پس سیناپسی را به دو نوع سریع اثر کوچک مولکول و نوروپپتیدهای درشت مولکول تقسیم بندی می کنند. پاسخ های سریع دستگاه عصبی مانند انتقال سیگنال های حسی به مغز و انتقال سیگنال های حرکتی به عضلات تحت تاثیر میانجی های کوچک مولکول صورت گرفته اما نوروپپتیدها مسئول تغییرات طولانی مدت نورونی مانند تغییرات طولانی مدت در تعداد گیرنده های نورونی، بازکردن یا بستن کانال های بوئی معین به مدت طولانی و تغییرات طولانی مدت در تعداد با اندازه سیناپس ها می باشند.
ناقلین سریع اثر کوچک مولکول در سیتوزول پایانه پیش سیناپسی ساخته شده و به طریق انتقال فعال به داخل وزیکول ها منتقل می شوند. زمانی که پتانسیل عمل به پایانه پیش سیناپسی می رسد، وزیکول های حاوی ماده میانجی محتویات خود را به داخل شکاف سیناپسی رها می کنند. در این حالت غشاء وزیکول با غشاء پایانه پیش سیناپسی جوش می خورد. سپس در عرض چند ثانیه تا چند دقیقه، قسمت وزیکولی غشاء مجددا وارد غشای پایانه پیش سیناپسی شده و به داخل سیتوزول وارد می شود.
ناقلین سریع اثر کوچک مولکول
برخی از مهم ترین ناقلین سریع اثر کوچک مولکول در زیر آمده اند:
1. استیل کولین (ACh): استیل کولین در پایانه پیش سیناپسی از استیل کوآنزیم A و کولین تحت تأثیر آنزيم کولین استیل ترانسفراز ساخته می شود و وارد وزیکول ها می گردد. آزاد شدن استیل کولین به شکاف سیناپسی سبب می شود تا آنزيم کولین استراز موجود در این قسمت، آن را به استیل و کولین تجزیه نماید. کولین حاصله به طور فعال به درون پایانه منتقل می شود تا مجددا برای ساخت استیل کولین به کار برده شود. استیل کولین بیشتر از قسمت های زیر ترشح می گردد ۱) پایانه های سلول های بزرگ هرمی در قشر حرکتی ۲) انواع مختلف نورون های موجود در گانگلیون قاعده ای، ۳) نورونهای حرکتی عصب دهنده به عضلات اسکلتی، ۴) نورون های پیش عقدهای دستگاه عصبی خودمختار، ۵) نورون های پس عقدهای دستگاه عصبی پاراسمپاتیک. بسته به اینکه استیل کولین به کدام نوع از گیرنده های خود متصل شود، می تواند اثرات تحریکی یا مهاری ایجاد کند.
2. نوراپی نفرین (NE): نوراپی نفرین بیشتر از هسته نیلی یا لوکوس سرولئوس پل مغزی ترشح می شود و در تنظیم فعالیت کلی مغز و خلق فرد کمک کننده است بسیاری از نورون های پس عقدهای سمپاتیک نیز NE را ترشح می کنند. اكثر اثرات NE در بدن تحریکی بوده اما در نواحی اندکی، گیرنده های مهاری را فعال می کند.
3. دوپامین (۱۸): از ماده سیاه (Substantia nigra) در عقده های قاعده ای ترشح شده و اثر مهاری دارد.
4. گلیسین: عمدتا در سیناپس های نخاع ترشح می شود و اثر آن مهاری می باشد.
5. گاما آمینوبوتیریک اسید (GABA): یک میانجی مهاری عمده در نخاع، مخچه، عقده های قاعدهای و قشر مغز می باشد.
6. گلوتامات (Glu): از پایانه های پیش سیناپسی بسیاری از مسیرهای حسی ورودی به دستگاه عصبی مرکزی و نیز بسیاری از نواحی قشر مخ ترشح می شود و همیشه به صورت تحریکی عمل میکند.
7. سروتونین (HT-5): سروتونین به طور عمده از سجاف میانی (median raphe) در ساقه مغز ترشح شده و در تنظيم خلق و تنظیم خواب و بیداری نقش دارد. اثرات سروتونین عمدتا مهاری می باشند.
8. اکسید نیتریک (NO): N0 در نواحی مغزی مسئول رفتارهای طولانی مدت و حافظه ترشح می گردد، تفاوت NO با دیگر میانجی های ریز مولکول در این است که این ماده در پایانه پیش سیناپسی ذخیره نمی شود بلکه به صورت آنی ساخته شده و در شکاف سیناپسی ترشح می گردد. اثر NO بر پایانه پس سیناپسی عمدتا تغییر فعالیت متابولیسمی داخل سلول است.
میانجی های درشت مولکول (نوروپپتیدها)
نوروپپتیدها عمدتا در جسم سلولی نورون ساخته شده، در شبکه اندوپلاسمی پردازش شده و پس از بسته بندی در دستگاه گلژی همراه با جریان اکسونی به صورت بسیار آهسته به انتهای پایانه پیش سیناپسی می رسند و در آنجا در پاسخ به پتانسیل عمل، محتویات خود را رها می سازند. نوروپپتیدها برخلاف میانجی های ریز مولكول سبب تغییرات طولانی مدت در نورون پس سیناپسی می گردند.
وقایع الکتریکی نورون
پتانسیل استراحت غشای نورون در حد ۶۵- میلی ولت می باشد. غلظت یون سدیم در خارج غشاء بیشتر از داخل و غلظت یون پتاسیم در داخل بیشتر از خارج است. علت این امر وجود پمپ سدیم - پتاسيم ATPase می باشد که در ازای بیرون ریختن یون سدیم از سلول، یون پتاسیم را وارد سلول می کند. غلظت یون کلر در خارج غشاء بیشتر از داخل آن است که احتمال وجود پمپ ضعیفی را برای کلر مطرح می کند.
پتانسیلی که با عبور یک یون مشخص به طور کامل مقابله می کند، پتانسیل نرنست (Nernst) برای آن یون نامیده می-شود. با محاسبه پتانسیل نرنست، این رقم برای سدیم ۶۱+ به دست می آید اما پتانسیل واقعی غشاء ۶۵- میلی ولت است. بنابراین پمپ سدیم، آن دسته از یون های سدیم را که به درون سلول نشت می کنند، فورا به خارج از سلول پمپ می کند تا پتانسیل ۶۵- میلی ولت درون سلول را حفظ کند. پتانسیل نرنست برای پتاسیم ۸۶- میلی ولت است و برای این که پتانسیل غشا به این حد برسد، یون پتاسیم باید از سلول خارج شوند اما پمپ پتاسیم با این خروج مقابله می کند. پتانسیل نرنست برای یون کلر ۷۰- میلی ولت است. بنابراین کلر در حالت عادی تمایل دارد وارد سلول شود.
پتانسیل پس سیناپسی تحریکی
زمانی که یک ماده ناقل تحریکی بر نورون پس سیناپسی عمل می کند سبب بازشدن کانال های سدیمی و یا کلسیمی غشای نورون پس سیناپسی می گردد. هجوم یون مثبت سدیم و یا کلسیم به داخل سلول، پتانسیل استراحت غشاء را کاهش داده (به عنوان مثال از ۶۵- به ۴۵) و تمایل آن را به تحریک شدن افزایش می دهد. افزایش مثبت در ولتاژ (که در واقع یک مقدار منفی کمتر است)، پتانسیل پس سیناپسی تحریکی (EPSP) نام دارد، زیرا اگر پتانسیل به حد کافی در جهت مثبت افزایش یابد، تولید پتانسیل عمل در نورون پس سیناپسی تحریک می گردد.
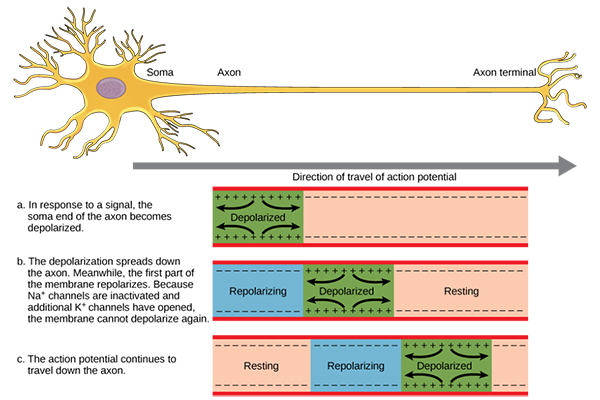
تولید پتانسیل عمل در نورون
زمانی که EPSP به حد کافی در جهت مثبت افزایش یافته و تا آستانه ایجاد پتانسیل عمل پیش برود، پتانسیل عمل در قسمت ابتدایی آکسون (تپه آکسونی) در محلی که آکسون، جسم سلولی نورون را ترک می کند، تولید می شود. علت اینکه پتانسیل عمل نورون تنها در این بخش از نورون تولید می شود این است که تعداد کانال های سدیمی وابسته به ولتاژ در این قسمت از نورون بیش از بقیه قسمت های نورون می باشد.
پتانسیل پس سیناپسی مهاری
زمانی که یک ماده ناقل مهاری بر غشاء نورون پس سیناپسی اثر می کند، سبب باز شدن کانال های کلری و یا پتاسیمی می گردد. از آنجا که پتانسیل نرنست برای یون کلر ۷۰- میلی ولت می باشد، باز شدن کانال های کلری با افزایش ورود کلر به داخل سلول می تواند پتانسیل داخل سلولی را از ۶۵- تا ۷۰- میلی ولت برساند. باز شدن کانال های پتاسیمی نیز با افزایش دادن خروج یون های مثبت پتاسیم از سلول سبب منفي ترشدن بیشتر پتانسیل استراحت غشای نورون می گردد. بنابراین افزایش در مقدار منفی بودن پتانسیل غشاء به بیش از حد پتانسیل استراحت غشا در حالت طبیعی که موجب کاهش تحریک پذیری سلول می گردد، پتانسیل پس سیناپسی مهاری (EPSP) نام دارد.
مهار پیش سیناپسی
برخلاف مهار غشاء نورون پس سیناپسی که قبلا گفته شد، نوع دیگری از مهار تنها در نورون پیش سیناپسی رخ می دهد و سبب کاهش آزادسازی ماده میانجی از این پایانه ها می گردد. در اکثر موارد، ماده ناقل مهاری GABA است. این ماده سبب افزایش نفوذپذیری غشاء پیش سیناپسی به یون کلر و افزایش ورود کلر به آن می گردد. بارهای منفی کلر، انتقال سیناپسی را مهار می کنند زیرا آنها قسمت اعظم اثر تحریکی یون های مثبت سدیم که با رسیدن پتانسیل عمل به درون رشته انتهایی وارد شده اند را خنثی می کنند.
جمع فضایی در نورون ها
اگر یک پایانه پیش سیناپسی، یک میانجی تحریکی یا مهاری را آزاد سازد و این میانجی بر نورون پس سینایسی اثر کند، تنها می تواند نیم تا ۱ میلی ولت پتانسیل منفی غشا را کاهش یا افزایش دهد، در حالی که برای اینکه پتانسیل غشاء به حد آستانه تولید پتانسیل عمل برسد باید تا حد ۱۰ الی ۲۰ میلی ولت کاهش یابد. اگر چندین پایانه پیش سیناپسی مواد میانجی خود را به طور همزمان بر روی پایانه پس سیناپسی تخلیه کنند، پتانسیل های پس سیناپسی تحریکی و مهاری آنها می توانند در جسم سلولی نورون با یکدیگر جمع شوند و اگر پتانسیل استراحت غشاء را به میزانی مثبت نمایند تا به حد تولید پتانسیل عمل برسد، پتانسیل عمل ایجاد می گردد اثر جمع همزمان پتانسیل های پس سیناپسی ناشی از فعال شدن چندین پایانه در ناحیه وسیع روی غشای نورون، جمع فضایی (Spatial summation) نامیده می شود.
جمع زمانی
اگر بلافاصله پس از ایجاد پاسخ تحریکی (EPSP) نورون پس سیناپسی که ناشی از آزاد شدن یک میانجی تحریکی از پایانه پیش سیناپسی است، پایانه پیش سیناپسی دوباره همان نورون پس سیناپسی را تحریک کند، پاسخ تحریکی دوم با پاسخ اولیه جمع می شود و EPSP حاصله افزایش بیشتری می یابد. بنابراین تخلیه های موفقیت آمیز پایانه پیش سیناپسی اگر با سرعت کافی رخ دهند می توانند با هم جمع شوند. این نوع تجمع، جمع زمانیtemporal) (summation نام دارد.
تسهیل نورون ها
اگر یک EPSP و یک IPSP با هم به یک نورون اعمال شوند، می توانند یکدیگر را خنثی کنند، اما زمانی که جمع پتانسیل های پس سیناپسی تحریکی بیش از جمع پتانسیل های پس سیناپسی مهاری باشد، گفته می شود که نورون تسهیل شده است. یعنی پتانسیل استراحت آن به حد آستانه تولید پتانسیل عمل نزدیک شده است. بنابراین یک محرک کوچک در یک نورون تسهیل شده می تواند سبب تولید پتانسیل عمل در آن گردد.
نقش دندریتها در تحریک نورونی
دندریت های نورونی در محدوده بسیار وسیعی از اطراف نورون منشعب شده اند و میدان فضایی بزرگی دارند که آنها را قادر می سازد تا سیگنال های تحریکی و مهاری را از نواحی وسیعی دریافت کنند. با وجود اینکه دندریت ها قادر به تولید پتانسیل عمل نیستند، می توانند پتانسیل های پس سیناپسی تحریکی و مهاری ایجاد شده در خود را به شیوه هدایت الكترو تونیک (گسترش جریان الکتریکی از طریق هدایت یون ها در مایعات موجود در دندریت ها) به جسم سلولی و حتی ابتدای اکسون منتقل کنند. البته پیش از آنکه پتانسیل های تحریکی بتوانند به جسم سلول ها برسند، مقدار زیادی از این پتانسیل از طریق غشاء نشت می کند و از دست می رود. کاهش پتانسیل غشا به علت انتقالي الكتروتونیک سیگنال در طول دندریت به سمت جسم سلولی، هدایت کاهش یابنده نامیده می شود.
خصوصیات ویژه انتقال سیناپسی
خستگی انتقال سیناپسی
تحریک و تخلیه مداوم سيناپس های تحریکی سبب می شود تا سرعت تخلیه سیناپس به تدریج کاهش یابد. به این حالت، خستگی سیناپسی اطلاق می شود. علت خستگی سیناپسی می تواند اتمام کامل یا نسبی ذخایر مواد ناقل در پایانه های پیش سیناپسی، غیرفعال شدن تدریجی گیرنده های غشای پس سیناپسی و تجمع غیرطبیعی یون ها درون نورون پس-سیناپسی باشد. خستگی سیناپسی سبب می شود تا از فعالیت بیش از حد نورون ها جلوگیری شود.
اثر اسیدوز و آلکالوز بر انتقال سیناپسی
به طور کلی، افزایش pH مایعات بدن (آلكالوز)، تحریک پذیری نورون ها را به شدت افزایش داده و در موارد شديد حتی می تواند سبب تشنجات مغزی گردد اما کاهش pH مایعات بدن (اسیدوز)، فعالیت مغزی را کاهش داده و در موارد شدید سبب اغما می شود.
هیپوکسی
کاهش میزان اکسیژن در دسترس نورون ها، می تواند سبب از بین رفتن کامل تحریک پذیری آن ها شود.
اثر داروها بر انتقال سیناپسی
موادی مانند کافئین، تئوفیلین و تئوبرومین که به ترتیب در قهوه، چای و کاکائو یافت می شوند، تحریک پذیری نورون ها را افزایش می دهند. استریکتین که فعالیت مواد ناقل مهاری را مهار می کند نیز بر فعالیت نورونی اثر تحریکی دارد. داروهای بیهوش کننده با افزایش دادن آستانه تحریک غشای نورون ها، موجب کاهش انتقال سیناپسی در دستگاه عصبی می گردند.
تأخير سيناپسی
از زمان تحریک نورون پیش سیناپسی تا رهایش ماده میانجی، رسیدن ماده میانجی به نورون پس سیناپسی و اعمال اثر بر نورون پس سیناپسی، زمانی در حدود نیم هزارم ثانیه صرف می شود. به ایم زمان، تاخیر سیناپسی اطلاق می شود.
- تهیه : محمدحسین فلاح، دانشجوی پزشکی دانشگاه علوم پزشکی ایران
- ویرایش: علیرضا نوری، دبیر کارگروه نوروفیزیولوژی شاخه دانشجویی نقشه برداری مغز ایران




نوشته های مرتبط